
いびきの音が気になって我慢がならないとか、人の咀嚼音がダメだとか、どうしても特定の音に嫌悪感を強く持ってしまう人がいます。こういう人にとっては「いびき」が凶器となりうるのです。いびきなんて本人がかきたくてかいているわけじゃないんだからどうしようもないじゃないかと、開き直る前に、その実態をよく見てみましょう。
イギリスニューキャッスル大学の神経科学研究所による研究発表がなされました。
「ミソフォニア(音嫌悪症)」と呼ばれる、特定の音に対し極端に嫌悪を示し、場合によっては神経症や不安、パニックを引き起こす症状の原因が、脳の構造にあるのかもしれないという報告です。
気になる音というのは、いびきだけではなく、くちゃくちゃ食べる音や、ズルズルと麺を吸い込む音だったり様々です。
これといった解消法はないのですが、ほかの雑音で気を紛らわすのが効果的だそうです。
長距離深夜バスの中でいびきとか寝息が気になっても、実は車の走行音のほうが大きいのです。それでも気になる音というのは耳についてしまうし、それが自分の気分を害していると思うだけで我慢できない大きさに聞こえます。
イヤホンの音を大きくするよりも、好きな映画やドラマなどの何度も聞いているセリフのシーンを少し小さい音で聞くようにすると、雑音に紛れるセリフに気持ちが集中するので、ほかの音を頭から追い出す効果があります。
ただラジオなどでは、聞こえづらい声を一生懸命聞こうとして、目がさえてしまいますので、映画などのよく知っているシーンで、聞こえづらくても脳が補える程度が寝るためにもいいのです。
集合住宅で、隣の音や上の階の音が気になるという方が多いのも、ミソフォニアの可能性があります。重症となると、ほかの人には聞こえない音まで、幻聴のように聞こえ、本人は非常に不快になります。場合によっては刑事事件に発展することがあります。
以下は英語論文を機械翻訳しているため、若干日本語が分かりづらいですが、ご紹介いたします。
http://www.sciencedirect.com/science/article/pii/S0960982216315305
ミソフォニア(Misophonia)の脳の基礎
ハイライト
•トリガー・サウンドは、ミソフォニアの前孤立部で誇張された反応を引き出す
•ミソフォニアでは、前部膵島(island of Reil):「大脳半球の外側溝の底にある大脳皮質の一部;自律機能,嗅覚きゅうかく,情動などをつかさどる.」の異常な機能的接続性がある
•自律神経反応の亢進は、ミソフォニアの前腸内括約筋によって媒介される
•ミソフォニアは相互作用の変化に関連している
概要
ミソフォニアは、他の人々が食べたり、飲んだり、噛んだり、呼吸したりするような日常的な音に反応して、強い負の感情(怒りと不安)を経験するという情動的な音声処理障害である[ 1-8 ]。これらの音の一般的な性質(しばしば「トリガー音」と呼ばれる)は、ミソフォニアを被害者およびその家族にとって壊滅的な障害にするが、根底にあるメカニズムについては何も知られていない。生理学的測定と連動した機能的および構造的MRIを用いて、ミソフォニック被験者は脳および身体において特定のトリガー音関連反応を示すことを示す。具体的には、fMRIは、発声された被験者において、トリガー音が、誇張された血液酸素レベル依存性(sensory processing)の知覚に重要な「顕著性(salience)」ネットワークの中枢である前部皮質皮質(anterior insular cortex :AIC)における応答(BOLD)ミソフォニックのトリガー音は、腹側前頭前野(vmPFC)、後頭皮質(PMC)、海馬および扁桃体を含む感情の処理および調節を担当する領域のネットワークと、AICとの間の異常な機能的接続と関連していた。トリガー・サウンドは、心拍数(HR)の上昇と皮膚反応(GSR)を測定した。アンケート分析によれば、ミソフォニックの被験者は、身体を異なる方法で知覚することができ、AICの異常な機能と一致して、対照よりも感覚的感受性が高いスコアを示した。最後に、脳構造測定は、ミソフォニア個人においてvmPFC内でより大きな髄鞘形成を示唆した。全体として、我々の結果は、ミソフォニアは、AICの異常な活性化および機能的接続性に基づいて異常な顕著性が特定の音に起因する障害であることを示している。
キーワード
ミソフォニア ;感情障害 ;fMRI ;機能的な接続性。自律的な応答 ;介入
結果と考察
fMRIデータは、20音色のミソフォンと22の年齢と性別が一致した対照群で取得され、トリガー音(食べること、呼吸する音などのミソフォニックな反応ではミソフォニックな反応を引き起こす)、不快な音両方のグループによって迷惑であると認識されるが、赤ちゃんの叫び声、悲鳴を上げる人物などのミソフォニック・ストレスを感じさせない)、中立的な音(例えば雨)を感じる。被験者は、それぞれの音を聞いた後、(1)どのように迷惑をかけるか(両方のグループ)、(2)効果的に典型的なミソフォニック反応(ミソフォニックグループのみ)を引き起こしたか、音が発生する環境にあること)、音は(制御グループのみ)であった。行動反応、電気的皮膚反応(GSR)および心拍数(HR)は、fMRIデータの取得中に取得された(パラダイム図の図1A 参照)。全脳構造のMRIデータは、髄鞘形成の含量、水分、および鉄分量を測定するためのマルチパラメータマップ(MPM)[ 9 ]として取得された。
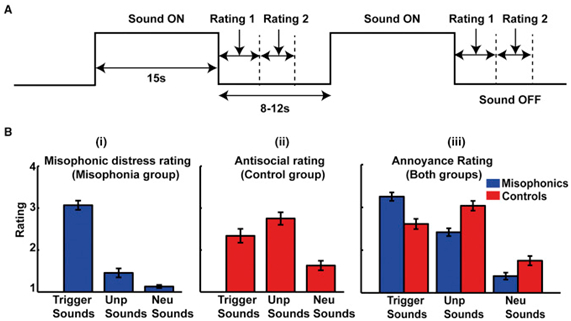
図1。
実験的パラダイムと主観的評価
(A)fMRIパラダイム:音を15秒間提示する標準的なブロックデザインを使用した。すべての音の後、被験者は、(1)音がどのように迷惑であったか、および(2)音がミソフォニック反応(ミソフォニア群)を引き起こすか、または反社会的であるか(対照群)であった。fMRIデータは、繰り返し時間(TR)が3.12秒で連続的に取得された。GSRおよびHRも実験を通してモニターした。
(B)主観評価:(i)ミソフォニックグループによる3種類の音のミソフォニック遭難評価。(ii)音の反社会的評価(対照被験者)。(iii)両方のグループによる音の苛立ち評価。不快感のある被験者は、不快な音(p <0.001)および中立的な音(p <0.001)と比較して、より大きなミソフォニック反応を引き起こすとトリガー音を評価した。ミソフォニックの被験者による不快な音は、迷惑であると認識され(中性音と比較してp <0.001)、一般的な迷惑とミソフォニック反応との間の解離を実証した。体感に関する主観的スコアについては、図S4も参照のこと。データは平均(±SEM)として表す。
行動データ(図 1B)は、トリガーサウンドがミソフォニックの被験者においてミソフォニック苦痛を引き起こした一方、不愉快なサウンドは、迷惑ではあるものの、ミソフォニック反応をもたらさなかったことを示した。ミソフォニックグループによるトリガー音のミソフォニック苦悩評価と、対照グループによる不快音の苛酷度格付けとの間に差はなかった。しかし、2つのグループは音を評価しながら異なる主観的尺度を使用した可能性が高い。グループ(2つのレベル)と音のタイプ(3つのカテゴリ)を因子とする一般線形モデル(GLM)[ 10 ]を用いたfMRIデータのランダム効果分析は、前部皮質皮質(AIC)における相互作用を左右対称に示した(図 2A;さらに地域が表S1)。さらなる分析は、AICにおける相互作用がサウンドをトリガーに応答して対象を制御するために比べミソフォニック被験者においてより大きな活性化によるものであったことが示された(参照、図2のB及び図S1を確認するためのプロット;参照S2図)。ミソフォニックとコントロールの被験者間の有意な活性化の差異は、不快な音や中立的な音には生じなかった。で確認プロットに示すように、左右のAICの両方における活性が、ミソフォニックグループ内ミソフォニック苦痛の主観的評価に伴って直線的に変化し、図2証拠のC. A大体[ 11]は、AICが怒りを含む感情に関連する主観的感情に関与することを示唆している。機能的には、AICは、顕著性ネットワーク〔のキーノードであることが知られている12 ]、検出および個人の行動関連性と意味のある刺激に向かって注意を向けるための固有の大規模な脳ネットワーク。AICの特有の活動亢進は、発声者がこれらの音に異常に高い顕著性を割り当てるという仮説を支持する。
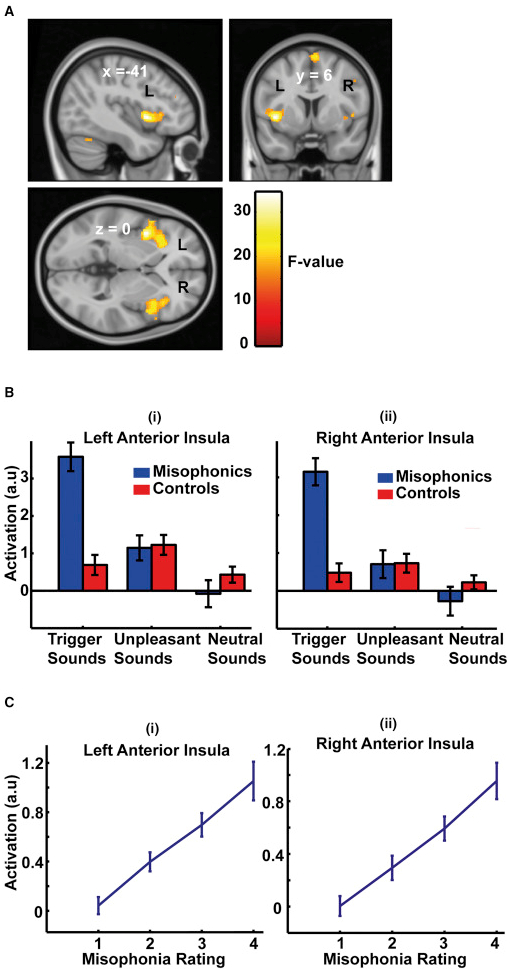
図2 グループレベルのランダム効果GLMによるfMRIデータの解析
GLMは、グループ(2つのレベル)と音のタイプ(3つのレベル)を要因とする階乗的な設計としてモデル化されました。
(A)2つの因子(群および聴覚のタイプ)の間の重大な相互作用のために標準的なMNI-152テンプレート脳にオーバーレイされた統計的パラメータマップ(SPM)であり、p = 0.05家族全体の誤差(FWE) 。この効果はMNI座標(-41,6,0)で最大値を有するAIC(両側)において最大である。
(B)AICのクラスターで平均した活動の確認プロット(図S1およびS2および表S1も参照)は、対話効果が、対照と比較して、ミソフォニック対象におけるトリガー音の活動が高いことによって引き起こされたことを示している。
(C)ミソフォニック科目におけるミソフォニック・レーティングを用いたAICにおける活動の確認プロット。
(B)および(C)のデータは平均(±SEM)を示す。
ミソフォニック参加者のトリガー音を区別する重要な領域としてAICを特定したので、ミソフォニアに特有のネットワークレベルの変更があるかどうかを確認するために、刺激依存の接続プロファイルを探求しました。左AICをシード領域として使用して、2つの群における刺激依存性の接続性を分析した。腹側前頭前野(vmPFC)、後内皮層(PMC;後部帯状疱疹および後退性皮質)、海馬および扁桃体を含む脳領域のネットワークにおいて、ミソフォニック対象に対するAICのより大きな機能的連結性が観察された(図3A)。この増加した機能的な接続性は、トリガー音に特有のものであった。不快な音に関しては、接続性に大きな違いは見られなかった。重要なことに、同じ音に対する2つのグループ間の機能的接続パターンは、量的にも質的にも異なっていただけでなく、vmPFCへの接続性はミソフォニックの被験者における(ニュートラルな音の接続性に関して)音のセットは否定的です。右AICの機能的接続性の分析はまた、vmPFCおよびPMCに対するトリガー音に特異的な増加した結合性を示した(図S3A ;扁桃体および海馬への機能的連結も観察されたが、わずかに緩和した閾値であった)。vmPFCとPMCは、デフォルトモードネットワーク(DMN)[ 13 ](参照図S3被験者が内部向け思考および記憶の検索に従事しているときに活性化されるDMNとAICの機能的接続ネットワークとの間のオーバーラップのためにB)を、[ 14 ]及び非活性化されます注目は外部刺激に向けられています。AICとDMNのより大きな結合は、発声音を聞くとミソフォニックな被験者がAICをDMNから「解放」することができないことを示唆している。これは最近の研究[ 15多変量パターン分類を用いて、vmPFCおよびPMCにおける活性のパターンが、異なるタイプの感情を区別するのに最も有益であることを示した。AICとvmPFCとPMCとの間で、ミソフォニックと同じ音に対する制御が明確に異なることは、これらの領域が、2つのグループのトリガー音に対して異なる感情反応を起こす上で重要な役割を果たすことを示唆しています。したがって、この非定形的な機能的接続性は、AICの異常な活性化、およびミソフォニックグループによって音を誘発するように割り当てられた異常な顕著性の根底にある可能性がある。
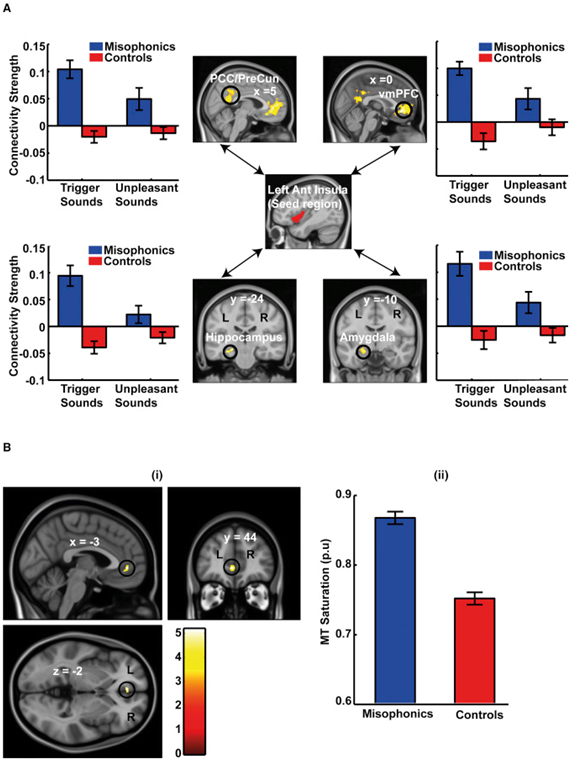
図3 機能的接続性と構造データ解析
(A)左AICをシード領域とし、脳のすべてのボクセルに対する機能的連結性を分析した。この図は、ミソフォニックな被験者(コントロールと比較して)におけるトリガーサウンド(ニュートラルサウンドと比較して)のより大きな接続性を示す脳領域を示しています。閾値を超えて生存する4つの領域は、(1)PMC(後部帯状皮質[PCC] /プレグネナス)、(2)vmPFC、(3)海馬、および(4)扁桃体である。各地域の棒グラフは、中立的な音に関するトリガーと不快な音の接続性の確認プロットを示しています。表示された接続強度は、p <0.05でクラスター閾値に設定され、クラスター形成しきい値はp <0.001になります(図 A-3の右側のAICの機能接続性および接続ネットワークとデフォルトモードネットワークのオーバーラップ)。
(B)失調症の脳構造変化。ミソフォニック被験者は、vmPFCのコントロールと比較して、より高い髄鞘形成を反映するより高いMT飽和を示す。複数の比較のために補正した場合(発声するためにミソフォニックでより高い機能的結合性を示す脳領域、すなわち、(A)のシード領域AICとともに示される機能的ネットワーク)で補正した場合、15個のボクセルのvmPFCが最大値(-3、44、-2)は補正後も残る。図の表示目的のために、補正されていないp <0.001の閾値が使用される。pu、パーセント単位。
棒グラフのデータは平均(±SEM)を示す。
ミソフォニアの症状は人生の早い段階で開始する(発症年齢は約12歳であると早くも5年もすることができます意味[ので1 ])、我々はまた、対照と比較してミソフォニック被験者における脳の構造的な違いがあるだろうと予測しました。我々は、脳灰白質における髄鞘形成を反映する磁化転移(MT)飽和の全脳構造マップを作成した。有意性試験のために、私たちは、シード領域と共に対照と比較して、ミソフォニックにおけるAICとのより高い機能的結合性を示す脳領域への探索を制限した。構造地図の分析は、ミソフォニック対象がMT飽和を変化させたことを示し、これはvmPFCの灰白質における有意に高いミエリン化と一致する(図 3B)。この変化は、ミソフォニックの被験者で観察されたvmPFCへの変化した機能的接続のための可能な構造的基礎を示唆している。
脳の機能的および構造的変化を同定した後、我々は次に、脳における身体およびその駆動源の生理学的反応を決定した。我々はGSRとHRを測定し、被験者はMRIスキャナーで3セットの音を聞いた。トリガー音は、ミソフォニックな被験者において、対照被験者よりも大きなGSR応答およびHR応答を誘発した(図4A)。生理的反応は健全な呈示の持続期間を通じて持続し、不快なニュートラルな音のための2つのグループ間のGSR応答またはHR応答に差異がなく、音を誘発することに特有であった。私たちが観察したトリガー特有の自律神経反応の高まりは、トリガー音の環境から逃れる発声者の強い傾向と一致している[ 1] 。 2 ]逃げることができなければ強い不安と怒りを経験する(戦闘/飛行の応答)。
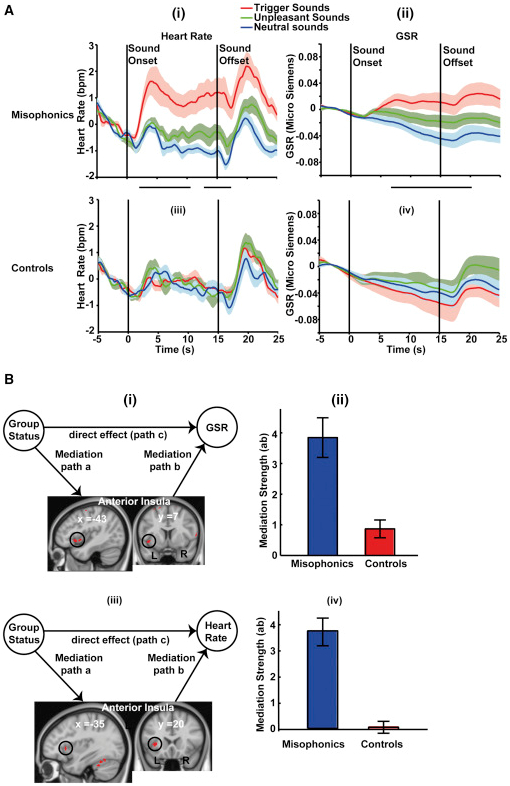
図4。脳領域による精神生理学的反応と仲介
(A)ミソフォニックおよびコントロール被験者のHRおよびGSR。ミソフォニックの被験者では、トリガー音がHRおよびGSRの持続的な増加をもたらす。GSRおよびHRの統計分析は、fMRI分析と同様に、2×3のANOVAを用いて時々実行された。HR時系列では、因子間の相互作用は、発症後2.4〜10.4秒、その後12.4〜17秒に有意であった。GSRの時系列では、発症後7〜21.4秒の有意な相互作用が観察された(GSRとHRが有意に異なる時点をパネル間に黒色の水平バーで示す)。HRおよびGSR時系列の両方は、p <0.05でクラスター閾値を有し、クラスター形成閾値はp <0.05であった。ポストホック比較は、HRとGSRの両方における相互作用効果が、ミソフォニックな被験者のトリガー音に対するより高い応答によって引き起こされることを示した。2つのグループの間には、不快でニュートラルな音に対する反応に違いはなかった。bpm、1分あたりのビート。
(B)調音分析は、発声音を発するために、対照と比較して、発声された被験者においてどの脳領域がHRおよびGSRの増加を仲介するかを決定する。入力Xはカテゴリーベクトル(ミソフォニックの場合は+1、コントロールの場合は-1)であり、応答ベクトルYはHR / GSRの平均的な増加を含んでいる(ニュートラルサウンドと比較して)全脳の一段階仲介分析が使用された)を各被験者のトリガー音のトライアルで聴く。仲介変数Mは、中立的な音と比較されるトリガー音のベータ値(SPMを使用して決定される)です。(i)左AICはGSRの変化を媒介する。(ii)AICのクラスターを平均した2つのグループのGSRのメディエーション強度の確認プロット。(iii)AICは、ミソフォニックにおける高められたHRを仲介する。(iv)AICのクラスターを平均した、2つのグループのHRの調停強度の確認プロット。
データは平均(±SEM; Aの陰影区域およびBのエラーバー)として表される。
ミソフォニアにおけるこれらの自律神経反応の脳の原因は何ですか?これに答えるために、我々は、変数X(グループメンバーシップ、すなわちミソフォニックまたはコントロール)からY(GSRまたはHR)への関係が第3の変数によって説明(媒介)できるかどうかをテストすることを目的とする仲介分析[ 16 ] M(脳活性化)。重大な仲介は、グループメンバーシップ(X)によって説明される以上に、観察されるGSR / HR(Y)を仲介することができるY(XからMからY)への間接的経路があり、 。我々はGSRとHRのために全脳仲介分析を別々に実施した。本発明者らは、AICにおける活性が、ミソフォニック対象におけるGSRおよびHRの上昇(図 4B)の両方を媒介することを見出した。
過去10年間で、インタオプション(内部身体状態の認識)が刺激に関連する感情の顕著さと経験に影響を与える可能性があるという認識が高まっている[ 17-20 ]。興味深いことに、AICは身体からの内臓の内臓入力を外部の感覚入力と統合する重要な脳構造である。これに伴い、AICの非定型的な介入と活性化は、多くの社会的情動障害の根底にあることが示されている[ 21,22 ]。最近、インターレセプションのモデルとして、予測に基づく階層的ベイズ推定を拡張への関心が高まってきた、19 23]。このモデルでは、インタセプトには、ボトムアップのインターセプト信号とその原因の事前の信念(予測)を組み合わせることで、インターセプト信号の原因を推測することが含まれます。このマルチレベルで階層的に構成された推論スキームでは、AICは階層の最上位にあり、身体の全体的な状態を推測することが示唆されている[ 24 ]。身体意識アンケート[ 25 ]を用いた身体知覚に関する主観的信念の評価は、ミソフォニクスが、ミソフォニックにおける変化した知覚感受性と互換性のある内部感覚(図S4)のより大きな意識を報告することを示した[ 22 ]。身体状態を表すAICの役割を考えると、アンケートデータはまた、ミソフォニアにおける異常なAIC機能と一致している。
結論
全体的に、我々のデータは、ミソフォニックの場合、トリガー音がAICの活動亢進およびこの領域の内側前面、内側頭頂および側頭領域との異常な機能的連結を引き起こすことを示している。AICとの異常な機能的接続性を示す内側前頭皮質における異常髄鞘形成が存在すること; 異常な神経応答が、ミソフォニック体験に伴う感情的な発色および生理的な覚醒を媒介することを示唆している。一緒になって、私たちのデータは、内的身体状態の非定型的な知覚と相まって、それ以外の無害な音に起因する異常な顕著性は、ミソフォニアの根底にあることを示唆している。利用可能なデータでは、ミソフォニアが非定型的な相互受話の原因であるのかそれとも結果であるのかを判断することは不可能であり、両者の関係を描写するためにさらなる作業が必要である。
ミソフォニアは、障害の神経学的または精神医学的分類を特徴としない。被害者は、これが引き起こす恐れの恐怖のためにそれを報告せず、医師は一般にその障害に気づいていない。この研究は、この悪性疾患を分類し治療するための継続的な努力を導く、行動、自律神経反応、および脳活動および構造の変化に基づく明確な表現型を定義する。
著者寄稿
SK、OT-H。、WS、JSW、およびTDGは実験を設計した。SKおよびOT-H。データを収集した。SKは、WS、JSW、MFC、TDGの助けを借りてデータを分析しました。SK、OT-H、WS、JSW、MFC、MA、TEC、PEG、D.-EB、およびTDGはこの論文を書いた。TDGは作業の全側面を監督しました。
謝辞
TDGは、このプロジェクトの財政的支援のためにウェルカム・トラストに感謝したい(付与WT091681MAおよびWT106964)。JSWは、MB / PhD卒業生のためのウェルカム・トラスト・ポスドク研修フェローシップ(グラント095939)を保持しています。ウェルカム・トラスト・センターは、ウェルカム・トラスト(助成金091593)からのコア・ファンドにより支援されています。この調査はUCL倫理委員会によって承認された
References
1
S. Kumar, O. Hancock, T. Cope, W. Sedley, J. Winston, T.D. Griffiths
Misophonia: a disorder of emotion processing of sounds
J. Neurol. Neurosurg. Psychiatry, 85 (2014), p. e3
CrossRef | View Record in Scopus | Citing articles (2)
2
M. Edelstein, D. Brang, R. Rouw, V.S. Ramachandran
Misophonia: physiological investigations and case descriptions
Front. Hum. Neurosci., 7 (2013), p. 296
View Record in Scopus | Citing articles (1)
3
A. Schröder, N. Vulink, D. Denys
Misophonia: diagnostic criteria for a new psychiatric disorder
PLoS ONE, 8 (2013), p. e54706
CrossRef
4
A. Schröder, R. van Diepen, A. Mazaheri, D. Petropoulos-Petalas, V. Soto de Amesti, N. Vulink, D. Denys
Diminished N1 auditory evoked potentials to oddball stimuli in misophonia patients
Front. Behav. Neurosci., 8 (2014), p. 123
5
M.M. Jastreboff, P.J. Jastreboff
Components of decreased sound tolerance: hyperacusis, misophonia, phonophobia
ITHS News Lett., 2 (2001), pp. 5–7
View Record in Scopus | Citing articles (8)
6
J.T. Krauthamer
Sound-Rage: A Primer of the Neurobiology and Psychology of a Little Known Anger Disorder
Chalcedony Press (2013)
7
G. Hadjipavlou, S. Baer, A. Lau, A. Howard
Selective sound intolerance and emotional distress: what every clinician should hear
Psychosom. Med., 70 (2008), pp. 739–740
CrossRef | View Record in Scopus | Citing articles (17)
8
G.M. Ferreira, B.J. Harrison, L.F. Fontenelle
Hatred of sounds: misophonic disorder or just an underreported psychiatric symptom?
Ann. Clin. Psychiatry, 25 (2013), pp. 271–274
View Record in Scopus | Citing articles (8)
9
N. Weiskopf, J. Suckling, G. Williams, M.M. Correia, B. Inkster, R. Tait, C. Ooi, E.T. Bullmore, A. Lutti
Quantitative multi-parameter mapping of R1, PD(∗), MT, and R2(∗) at 3T: a multi-center validation
Front. Neurosci., 7 (2013), p. 95
10
K.J. Friston, A.P. Holmes, K.J. Worsley, J.P. Poline, C.D. Frith, R.S.J. Frackowiak
Statistical parametric maps in functional imaging: a general linear approach
Hum. Brain Mapp., 2 (1994), pp. 189–210
CrossRef | View Record in Scopus | Citing articles (6735)
11
A.D. Craig
How do you feel—now? The anterior insula and human awareness
Nat. Rev. Neurosci., 10 (2009), pp. 59–70
View Record in Scopus | Citing articles (2161)
12
W.W. Seeley, V. Menon, A.F. Schatzberg, J. Keller, G.H. Glover, H. Kenna, A.L. Reiss, M.D. Greicius
Dissociable intrinsic connectivity networks for salience processing and executive control
J. Neurosci., 27 (2007), pp. 2349–2356
CrossRef | View Record in Scopus | Citing articles (2023)
13
M.E. Raichle, A.M. MacLeod, A.Z. Snyder, W.J. Powers, D.A. Gusnard, G.L. Shulman
A default mode of brain function
Proc. Natl. Acad. Sci. USA, 98 (2001), pp. 676–682
CrossRef | View Record in Scopus | Citing articles (5500)
14
W. Huijbers, C.M. Pennartz, R. Cabeza, S.M. Daselaar
The hippocampus is coupled with the default network during memory retrieval but not during memory encoding
PLoS ONE, 6 (2011), p. e17463
CrossRef | View Record in Scopus | Citing articles (1)
15
H. Saarimäki, A. Gotsopoulos, I.P. Jääskeläinen, J. Lampinen, P. Vuilleumier, R. Hari, M. Sams, L. Nummenmaa
Discrete neural signatures of basic emotions
Cereb. Cortex, 26 (2016), pp. 2563–2573
CrossRef | View Record in Scopus | Citing articles (6)
16
T.D. Wager, C.E. Waugh, M. Lindquist, D.C. Noll, B.L. Fredrickson, S.F. Taylor
Brain mediators of cardiovascular responses to social threat: part I: reciprocal dorsal and ventral sub-regions of the medial prefrontal cortex and heart-rate reactivity
Neuroimage, 47 (2009), pp. 821–835
Article | PDF (1386 K) | View Record in Scopus | Citing articles (134)
17
A.D. Craig
Interoception and emotion: a neuroanatomical perspective
M. Lewis, J.M. Haviland-Jones, L.F. Barett (Eds.), Handbook of Emotions, Guilford Press (2007), pp. 272–290
18
S. Wiens
Interoception in emotional experience
Curr. Opin. Neurol., 18 (2005), pp. 442–447
View Record in Scopus | Citing articles (157)
19
A.K. Seth
Interoceptive inference, emotion, and the embodied self
Trends Cogn. Sci., 17 (2013), pp. 565–573
Article | PDF (1334 K) | View Record in Scopus | Citing articles (179)
20
S.N. Garfinkel, H.D. Critchley
Interoception, emotion and brain: new insights link internal physiology to social behaviour. Commentary on: “Anterior insular cortex mediates bodily sensibility and social anxiety” by Terasawa et al. (2012)
Soc. Cogn. Affect. Neurosci., 8 (2013), pp. 231–234
CrossRef | View Record in Scopus | Citing articles (56)
21
M.P. Paulus, M.B. Stein
Interoception in anxiety and depression
Brain Struct. Funct., 214 (2010), pp. 451–463
CrossRef | View Record in Scopus | Citing articles (130)
22
S.N. Garfinkel, C. Tiley, S. O’Keeffe, N.A. Harrison, A.K. Seth, H.D. Critchley
Discrepancies between dimensions of interoception in autism: implications for emotion and anxiety
Biol. Psychol., 114 (2016), pp. 117–126
Article | PDF (1541 K) | View Record in Scopus | Citing articles (8)
23
A.K. Seth, K.J. Friston
Active interoceptive inference and the emotional brain
Philos. Trans. R. Soc. Lond. B Biol. Sci., 371 (2016), p. 20160007
CrossRef
24
R. Smith, R.D. Lane
The neural basis of one’s own conscious and unconscious emotional states
Neurosci. Biobehav. Rev., 57 (2015), pp. 1–29
Article | PDF (4216 K) | CrossRef | View Record in Scopus | Citing articles (12)
25
L.C. Miller, R. Murphy, A.H. Buss
Consciousness of body: private and public
J. Pers. Soc. Psychol., 41 (1981), pp. 397–406
CrossRef | View Record in Scopus | Citing articles (227)
ストレス減で活力ある未来に貢献する、株式会社RUDDER。
特許出願済み。まぶしい!うるさい!寒い!を解消。安眠家具「Sleep Labo」国産家具の安心安全をお届けします。
うるさいいびき、止まらない、止められない。でも大丈夫。 いびきを解決する唯一の方法。
お求めのショップへは緑ボタンをクリック(BASE)

代金引換現金でのご購入は黄色ボタンをクリック